Halothermophilic Cas9 and dCas9 Interaction with Variety sgRNA In-silico
Corresponding email: lischer.kenny@ui.ac.id
Published at : 31 Jan 2025
Volume : IJtech
Vol 16, No 1 (2025)
DOI : https://doi.org/10.14716/ijtech.v16i1.7367
Lischer, K, Hayati, K, Fabian, MH, Laksmi, FA, Nugraha, Y, Rizqi, HD, Fauziyya, R, Sarmoko & Ali, MSM 2025, 'Halothermophilic Cas9 and dCas9 Interaction with Variety sgRNA in-silico', International Journal of Technology, vol. 16, no. 1, pp. 322-331
| Kenny Lischer | 1. Bioprocess Engineering, Department of Chemical Engineering, Faculty of Engineering, Universitas Indonesia, Depok, West Java, 16424, Indonesia 2. Research Centre of Biomedical Engineering, Departme |
| Kholisoh Hayati | Chemical Engineering, Department of Chemical Engineering, Faculty of Engineering, Universitas Indonesia, Depok, West Java, 16424, Indonesia |
| Muhammad Haykal Fabian | Bioprocess Engineering, Department of Chemical Engineering, Faculty of Engineering, Universitas Indonesia, Depok, West Java, 16424, Indonesia |
| Fina Amreta Laksmi | National Research and Innovation Agency, Cibinong, West Java, 16915, Indonesia |
| Yudhi Nugraha | 1. Research Center for Molecular Biology Eijkman BRIN, National Research and Innovation Agency, Cibinong, West Java, 16915, Indonesia 2. Department of Molecular Medicine, University of Pavia, 27100, |
| Hamdan Dwi Rizqi | Department of Chemistry, Faculty of Science and Data Analytics, Institut Teknologi Sepuluh Nopember, Surabaya, Jawa Timur, 60111, Indonesia |
| Riri Fauziyya | Department of Pharmacy, Sumatera Institute of Technology, Lampung, 35365, Indonesia. |
| Sarmoko | Department of Pharmacy, Sumatera Institute of Technology, Lampung, 35365, Indonesia |
| Mohd Shukuri Mohamad Ali | Enzyme and Microbial Technology Research Center, Universiti Putra Malaysia, Serdang 43400, Selangor, Malaysia |

Industrial bioprocesses often require microorganisms that can adapt to specific conditions, such as halothermophilic bacteria capable of producing biosurfactants with various applications. Enhancing production yield is important, and one approach genetic modification. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) is a highly effective method for genetic editing; however, its use in halothermophilic bacteria remains limited. So far, optimal genetic editing with Cas9 has only been achieved in thermophilic bacteria unsuitable for high-salt environments. For determining optimal CRISPR involves analyzing the binding interaction between sgRNA (single guide RNA) and Cas9 using Molecular Docking in halothermophilic bacteria. This includes preparing the structures of Cas9 and sgRNA, simulating them with HDOCK software, and data analysis. Molecular Docking has advantages such as cost-effectiveness and time efficiency in designing the optimal sgRNA length. Optimization of sgRNA is achieved by varying the lengths of repeat, spacer, and tracrRNA, aiming for the lowest binding energy values and visually feasible designs. For Cas9 from the selected bacterium, Klebsiella pneumoniae, the optimal sgRNA design for Cas9 involves a spacer of 10 nt. In contrast, dCas9 involves a spacer of 10 nt, a repeat of 36 nt, and tracrRNA of 63 nt. The native temperature of the bacteria did not significantly affect the optimal sgRNA length.
Biosurfactant; CRISPR-Cas9 Optimization; Halothermophilic; Molecular docking; sgRNA Design
Nearly half of the global energy demand increase stems from rapidly developing countries, leading to a continuous rise in energy consumption each year. There are over 1.6 trillion barrels of oil reserves worldwide, but 67% of this total consists of residual crude oil in reservoirs that are difficult to process (Shibulal et al., 2014). One alternative for processing this residual oil involves the use of biosurfactants produced by bacteria, either in-situ or ex-situ, through the use of bioreactors (Geetha, Banat, and Joshi, 2018). Biosurfactants are also utilized in the cosmetic, pharmaceutical, food, and agricultural industries, serving as emulsifying agents, foaming agents, and solvents (Akbari et al., 2018). Owing to their diverse applications and environmentally friendly benefits, the biosurfactant market is projected to grow at a CAGR of 5.5% from 2023 to 2028 (Mordor Intelligence, n.d.). Halothermophilic bacteria, resistant to high temperatures and salinity, can optimally produce biosurfactants for commercial-scale production at temperatures above 40°C and are commonly found in hot springs, deep-sea hydrothermal vents, or high-salinity lakes, such as those in the Arabian Gulf (Roda and Bassam, 2017; Lischer et al., 2020; Elazzazy, Abdelmoneim and Almaghrabi, 2015). Halothermophilic bacteria have a wide range of potential applications in various industries, including dye decolorization, as well as in pharmaceuticals, cosmetics, and food. They are used in the production of extremophile lipase enzymes (Memarpoor-Yazdi et al., 2017) and biocatalyst production (Rodrigues et al., 2017). The optimization of biosurfactant production can be achieved through genetic engineering, such as inserting the rh1AB or rh1A genes to produce more biosurfactants or using mutagenesis to modify amino acid sequences (Jimoh et al., 2021). However, genetic engineering in halothermophilic bacteria remains limited due to the difficulties in gene editing and the lack of related research. The technology to do genetic engineering still needs to be developed. Current technology, such as TALEN, ZINC Finger, and current Cas9, cannot withstand the harsh environment where halothermopphilic bacteria grow. One genetic modification method is CRISPR/Cas9, which can cut DNA at specific locations and alter its sequence. CRISPR-Cas9 is a highly effective, efficient, and accurate genome-editing tool used across various scientific disciplines. Cas9 encodes a multidomain protein that targets and cuts DNA, while dCas9 binds to specific DNA segments without cutting, typically through modifications in the RuvC and HNH nuclease domains (Mengstie & Wondimu, 2021; Makarova et al., 2015). However, CRISPR/Cas9 is currently optimal at 37°C, which is less suitable for thermophilic bacteria (Mougiakos et al., 2017). Studies on CRISPR/Cas9 interactions in thermophilic bacteria have been conducted, identifying optimal spacer, repeat, and tracrRNA lengths for genetic modification (Pramayuditya, 2023; Angela, 2022). Cas9 application has been studied in halophilic bacteria in Halomonas spp. (Qin et al., 2018). However, research on halothermophilic bacteria is limited, with initial exploration using Single Amplified Genomes (SAGs) by (Grötzinger et al., 2018) To address the challenges of operating at high temperatures and salinity, further studies are needed on the potential of Cas9 and dCas9 in halothermophilic bacteria. This includes optimizing the lengths of spacers, repeats, and tracrRNA using Molecular Docking simulations to predict interactions and binding affinities (Karlson et al., 2021; Uppada et al., 2018). The findings from these analyses can inform the design of optimal sgRNAs for halothermophilic bacteria, followed by in vitro and in vivo studies to validate their accuracy and enhance biosurfactant production post-genetic modification. Most docking studies are conducted to observe the interactions between drugs and their targets (Sahlan et al., 2023; Husnawati et al., 2023; Sahlan, et al., 2020). However, this research differs in that it focuses on docking to analyze the interactions between gRNA and Cas9 as well as dCas9.
In general, the experiment is divided into three major tasks, with two of them able to be conducted in parallel: Protein Sequence Identification and 3D Visualization of Cas9/dCas9 and sgRNA, then Molecular Docking Simulation for each variant with respective variations.
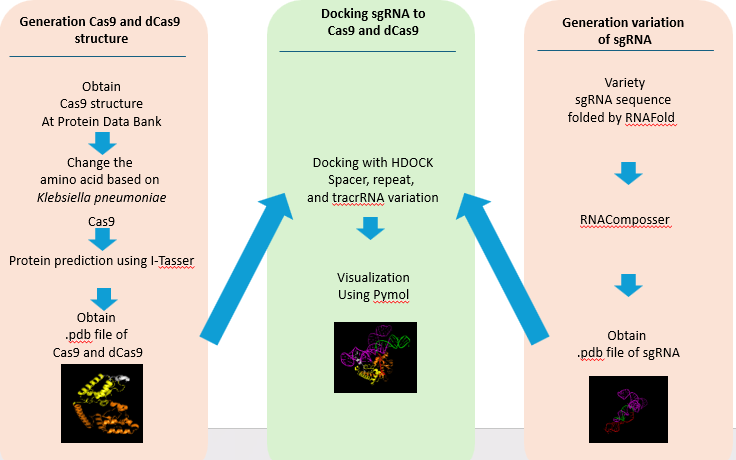
Figure 1 Scheme of the docking process
2.1. Preparation of Sequences and 3D Visualization of Cas9 and dCas9 Proteins from Halothermophilic Bacteria
The 3D structure visualization of Cas9 and dCas9 proteins can be performed after obtaining their respective protein sequences. In this study, Cas9 from the bacterium Klebsiella pneumoniae was used, sourced from the Protein Data Bank with the accession code MCI3999334.1. The protein sequence was obtained by accessing the NCBI website and searching for "type II CRISPR RNA-guided endonuclease Cas9 [target bacterium name]," in this study, we are using Klebsiella pneumoniae. The "Identical Protein Groups" database was selected, and the target protein sequence was accessed and downloaded in FASTA format.
Before visualizing, dCas9 was derived based on the study by Mougiakos et al. (2017) on ThermodCas9, which involves mutating the Cas9 sequence at amino acids 8 and 582. The 3D visualization was then performed using the cloud-based server I-TASSER, where the protein sequence file in FASTA format was uploaded, an email and password were entered to receive the results, an optional ID name was provided to distinguish the visualization results, and the I-TASSER tool was run (Zhou et al., 2022; Zheng et al., 2021; Yang and Zhang, 2015). The results were sent via email upon completion of the protein visualization.
2.2. Preparation of Sequences and 3D Visualization of sgRNA Variants
The sgRNA sequences used in this study are based on sgRNA previously utilized for thermophilic bacteria by Harrington et al. (2017), as detailed in supplementary data 3 of their journal. Each variant was derived as follows: the base spacer of 20 nt was directly obtained from the reference journal data. The 10 nt spacer was created by deleting 9 nt from the right side of the 19 nt spacer sequence: UGUAAGCGGAUGCCAUAUG. The 30 nt spacer was created by adding 9 nt to the left side of the 21 nt spacer sequence: GUCUGUAAGUCUGUAAGCGGAUGCCAUAUG. The repeat and tracrRNA sequences were directly taken from the journal's existing data. The folding of repeat and tracrRNA was performed using RNAfold website, and 3D visualization of sgRNA was carried out using RNAComposer (Sarzynska et al., 2023; Zuker, 2023).
2.3. Molecular Docking Simulation of sgRNA-Cas9/dCas9 Complex
The molecular docking process was performed using HDOCK(Yan et al., 2020), following these steps: The 3D structure pdb data of Cas9 from halothermophilic bacteria submitted as the receptor in “Input Receptor Molecule”. The 3D structure pdb data of sgRNA also submitted as the ligand in “Input Ligand Molecule”. The interaction site residues of sgRNA within Cas9 were manually entered through the “Advanced Options (Optionals)” > Receptor Binding Site Residue(s) section. The residues were entered in the format M: A, where “M” represents the residue number of the protein interaction site and “A” is the protein chain. The interaction sites are based on a study done by Sun et al. (2019). A name was given to the file, and a personal email address was provided so that the docking results could be sent directly to the specified address upon completion. The process was initiated by clicking submit to start the molecular docking. The results will include the top 10 models in 3D, along with binding affinity values (docking score), confidence score, ligand RMSD, and interface residues. These docking results could be downloaded by clicking “All the results in a package.”
2.4. Visualization of sgRNA-Cas9/dCas9 Interaction in Halothermophilic Bacteria
The sgRNA-Cas9 complex formed from the HDOCK simulation can be visualized using PyMOL or directly viewed on the website. Using PyMOL, the docking results must be in chain B format before visualization can be performed. The steps for visualization are as follows: Open the PyMOL application and select File > Open > the pdb file from the simulation. Type the command “alter lig_(x).pdb, chain=’B’” and save the chain B file by selecting File > Export Molecule > Save. For analysis with LigPlot+, open the application and select Open > PDB Files > Browse... Choose DIMPLOT and fill in domain 1 with *A and domain 2 with *B, then click Run to view the visualization.
3.1. Cas9
Protein Alignment Result
The Cas9 protein from Klebsiella
pneumoniae was aligned with the well-characterized Cas9 protein from Geobacillus
kaustophilus, as described by
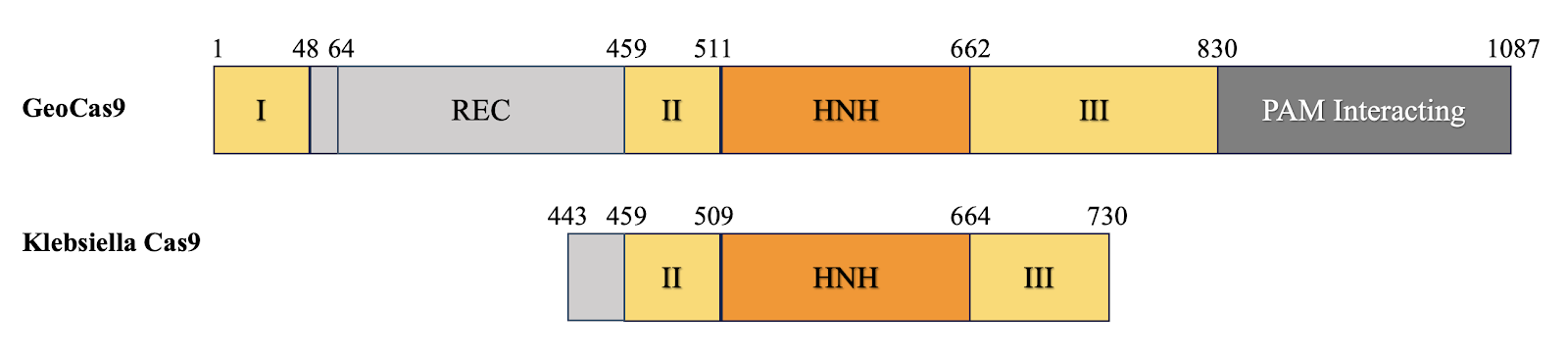
Figure 2 Visualization of Cas9 Domain
Cas9
from halothermopilic bacteria is significantly shorter (287 Amino Acids))
compared to geobacillus (1087 Amino Acid). The visualization results indicate a
significant difference in the coverage of target domains between the two
species, highlighting the necessity for specific mutations in the HNH domain to
achieve the desired functionality of dCas9 in Klebsiella pneumoniae.
This finding is crucial for understanding the structural and functional
adaptations required for effective gene editing in halothermophilic bacteria.
3.2. Visualization
of 3 Dimension Structure of Cas9/dCas9 and sgRNA
The second sequence visualized was sourced from the same locations: Air Panas Jono, Bleduku, and Grobogan, Bleduku in Semarang, East Java. The sequence contains 289 amino acid pairs, as shown in Figure 3.
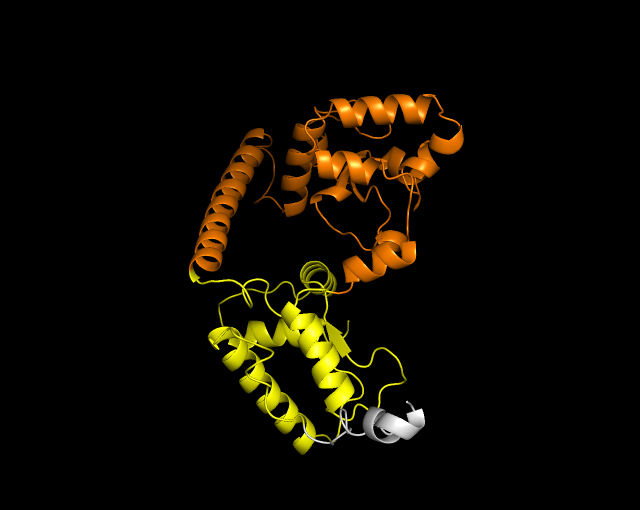
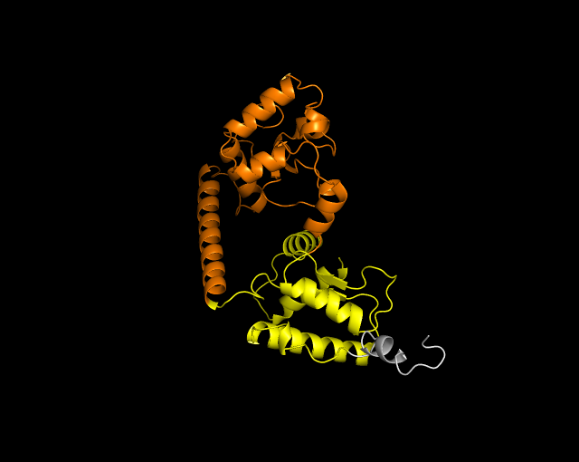
Figure 3 3-Dimension Structures (a) Cas9 and (b) dCas9 of Klebsiella pneumoniae
The colors in the structure represent specific areas: yellow for RuvC,
orange for HNH, and light gray for REC. Mutations were introduced exclusively
at positions A582F/A145F, which correspond to the HNH domain, due to the
limited Cas9 sequence data. The results show that the dCas9 fold is simpler,
with fewer alpha helices compared to Cas9. However, the structural similarity
is high, with an RMSD value of 0.542, indicating minor differences. This
suggests that the mutation affects functionality rather than structure.
For sgRNA, the spacer, repeat, and tracrRNA were folded separately since the spacer is expected to be linear. Initially, the construction of the 2D structure served as a bridge between the primary and tertiary structures, providing insights into how the sequence and base pairs form a functional 3D structure. It also confirmed that each sgRNA component forms the expected structure. Figure 4 shows the visualization of each sgRNA, with a red spacer, a repeat in green, and a tracrRNA in magenta. The visualization indicates that shorter nucleotide spacers form linear, unstructured regions. In contrast, longer nucleotides result in folds, which are undesirable as the spacer's function is to complement and bind the target DNA sequence for subsequent cutting.


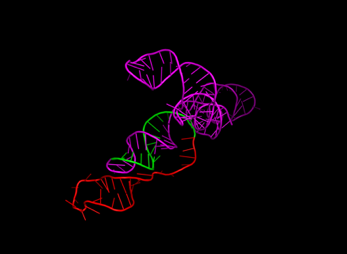
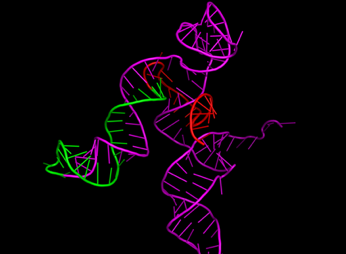
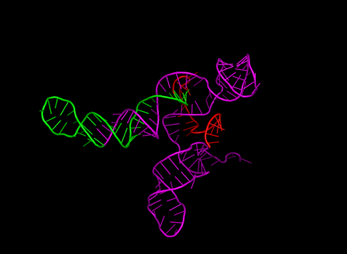
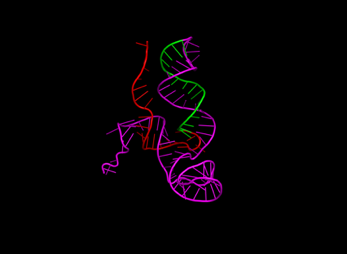

Figure 4 3-Dimensional Structures of
sgRNA with Variations: (a) Control, (b) Spacer 10 nt, (c) Spacer 30 nt, (d) Repeat 25 nt, (e) Repeat 36 nt, (f)
tracrRNA 63 nt, (g) tracrRNA 140 nt. (spacer in red, repeat in green, and tracrRNA in
magenta)
Furthermore, changes
in spacer length did not affect the structure or folding of the repeat and
tracrRNA regions. Notable differences were observed in the tracrRNA variants,
where different lengths altered the overall structure's orientation, indirectly
affecting the potential binding interactions.
3.3. Molecular
Docking Result of sgRNA-Cas9/dCas9 in Halothermophilic
After performing simulations
between sgRNA and Cas9, data consisting of the top 10 structures of the
sgRNA-Cas9/dCas9 complex were obtained. The structure selected was the top one
that most closely approached ideal conditions, where the spacer was positioned
closest to the RuvC and HNH domains. Docking was performed on Cas9/dCas9 from
Klebsiella pneumoniae, paired with sgRNA variants that incorporated changes in
spacer, repeat, and tracrRNA lengths. The goal was to observe their impact on
the formed complex and assess whether any variant formed a more stable complex
compared to the control. After identifying each variant of the spacer, repeat,
and tracrRNA for both Cas9 and dCas9, the best length of each variant was
selected by evaluating the lowest binding affinity values, highest confidence
scores, and the position where the spacer was linear and closest to RuvC and
HNH. For Cas9, the optimal sgRNA length was found to be a spacer of 10 nt, a
repeat of 16 nt, and a tracrRNA of 98 nt, with a binding affinity value of
-358.99 and a confidence score of 0.9849. Figure 5 (a) shows the linear spacer
(red) positioned between HNH (orange) and RuvC (yellow).
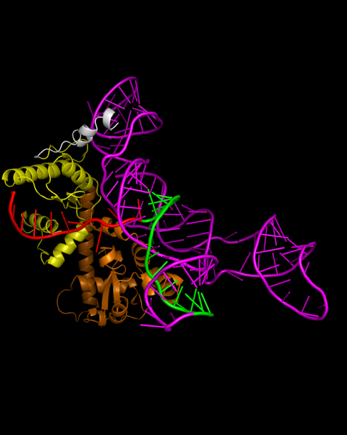
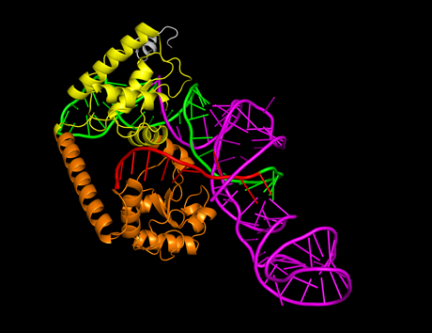
Figure 5 3-Dimension Structures of (a) Cas9 and (b) dCas9
The optimal sgRNA
design for dCas9 was a spacer of 10 nt, a repeat of 36 nt, and tracrRNA of 63
nt, with a binding affinity value of -354.28 and a confidence score of 0.9835.
Figure 4 (b) shows the 3D structure of the complex with the linear spacer (red)
positioned between HNH (orange) and RuvC (yellow) at a sufficiently close
distance. These results indicate that the specific lengths of spacer, repeat,
and tracrRNA significantly impact the binding affinity and overall structure of
the sgRNA-Cas9/dCas9 complex, providing insights into the optimal design for
effective gene editing in halothermophilic bacteria.
3.4. Influence of sgRNA Length on Binding Affinity
in Molecular Interaction
The visualization of intermolecular
interactions with varying spacer lengths showed that increasing the length of
the spacer in Cas9 does not significantly affect hydrogen bonding, whereas, for
dCas9, the opposite is true. This suggests that longer spacers can optimize the
positioning or flexibility of sgRNA, leading to better interactions with the
target DNA. This result is reflected in the binding affinity values/docking
scores in the two graphs in Figure 6, where the pattern tends to be linear with
the number of hydrogen bonds. From both graphs, it can be concluded that there
is a correlation between hydrogen bonds in the complex and binding affinity
values. An increase in the number of hydrogen bonds formed indicates enhanced
interaction and stability.
For the repeat region, there
is a correlation between nucleotide length and hydrogen bonding, especially in
dCas9. In Cas9, the most hydrogen bonds are found with a 25 nt repeat, while a
36 nt repeat shows a decrease due to the 3D structure folding away from Cas9.
This indicates that the length of the repeat affects the folding and
flexibility of sgRNA, thereby influencing its interaction with Cas9. The
binding affinity values/docking scores in Figure 7 show a similar pattern,
with a peak at 25 nt for Cas9 and a decrease at 36 nt. In the case of dCas9,
the variation in binding affinity values between 25 nt and 36 nt does not
correspond directly to the number of hydrogen bonds, possibly due to the flexibility
of both molecules and the arrangement of atoms within the folds. Comparing
these two parameters shows that repeat length significantly affects the number
of hydrogen bonds and docking scores, impacting the structural stability and
functional efficacy of the complex. The 25 nt repeat is most optimal for Cas9,
whereas the longest repeat is best for dCas9. However, the selection of sgRNA
length must consider the qualitative structure of the complex.
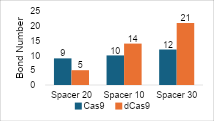
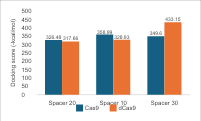
Figure 6 (a) Hydrogen Bond and (b) Intermolecular Binding Affinity on Spacer
Variations
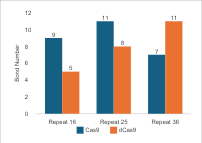
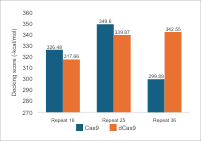
Figure 7 (a) Hydrogen Bond and (b) Intermolecular Binding Affinity on Repeat Variations
Regarding tracrRNA variations, the pattern in
Figure 8 shows that
in Cas9, longer tracrRNA results in more hydrogen bonds. However, in dCas9, the
98 nucleotide variation results in fewer bonds. Overall, this indicates that
longer tracrRNA increases the likelihood of forming bonds between Cas9 and
sgRNA. However, the binding affinity values for Cas9 decrease with longer
nucleotides, making the complex less effective in interactions. For dCas9, the
binding affinity values increase with longer tracrRNA, although this does not
always correlate with the number of hydrogen bonds formed. The results in
Figure 8
demonstrate that optimal binding is not solely dependent on the number of
hydrogen bonds but also on the configuration of sgRNA, particularly the
tracrRNA domain, which has the largest proportion in the complex.
Hydrogen bonds are
interactions between a hydrogen atom and an electronegative atom such as F, N,
or O. These bonds are stronger than Van der Waals forces but not as strong as
covalent bonds, making them ideal for biological interactions due to their stability
and flexibility
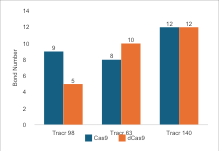
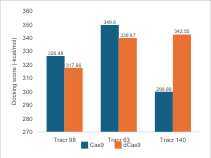
Figure 8 (a) Hydrogen Bond and (b) Intermolecular Binding Affinity on tracrRNA Variations
In molecular docking, hydrogen bonds are
calculated to predict the most stable and selective binding positions
The selected structures from the molecular
docking simulation were based on binding affinity values and visual validation
to ensure correct orientation and significant interactions
The appropriate design for sgRNA for Cas9 includes a spacer of 10 nt, a repeat of 16 nt, and tracrRNA of 98 nt. In contrast, for dCas9, the arrangement involves a spacer of 10 nt, a repeat of 36 nt, and a tracrRNA of 63 nt. Variations in spacer length do not significantly affect binding affinity values, whereas an increase in repeat length impacts binding affinity values. However, the binding affinity value decreases with increasing tracrRNA length in Cas9, which is opposite to what occurs in dCas9. The native temperature of the bacteria does not significantly affect the optimal sgRNA length for bacteria from that group. The results from in silico studies can be applied both in vivo and in vitro, especially in halothermophilic bacteria.
The author would like to extend sincere gratitude to the individuals and institutions for their invaluable assistance and support throughout this research: my supervisors for guiding me throughout the writing process, thesis examiners and the Department of Chemical Engineering, Faculty of Engineering, Universitas Indonesia, that participate in the process. This paper is supported by a fundamental research grant by Kemendibukristek, Indonesia Number: NKB-945/UN2.RST/HKP.05.00/2024.
Author Contributions
KL, FA, YN, do conceptual, analysis, supervise, and finalization draft of paper. HM draft first paper and analysis of the result. ACK, RWN do conceptual design and analysis. MSMA works on finalization of paper.
Conflict of Interest
The authors declare that there are no conflicts of interest
Akbari, S, Soudi, MR, Mousavi, SM & Hejazi, MA 2018, ‘Biosurfactants—a new frontier for social and environmental safety: a mini review’, Biotechnology Research and Innovation, vol. 2, no. 1, pp. 81-90, https://doi.org/10.1016/j.biori.2018.09.001
Angela, K 2022, Identifikasi bakteri berbasis lab-on-chip dari mata air panas cisolong banten, Final Thesis, Universitas Indonesia, Viewed (20 07 2024) (https://lib.ui.ac.id/detail?id=20523404&lokasi=lokal)
Arumsari, S, Handayani, ML & Nugroho, YS 2024, ‘Design of Specific and Efficient sgRNA for CRISPR/Cas9 System to Knockout Superoxide Dismutase 2 in Breast Cancer Stem Cells’, International Journal of Technology, vol. 15, no. 2, pp. 353-363, https://doi.org/10.14716/ijtech.v15i2.6680
Baker, EN & Htjabardt, RE 1984, ‘Hydrogen bonding in globular proteins’, Progress in Biophysics and Molecular Biology, vol. 44, no. 2, pp. 97-179, https://doi.org/10.1016/0079-6107(84)90007-5
Elazzazy, AM, Abdelmoneim, TS & Almaghrabi, OA 2015, ‘Isolation and characterization of biosurfactant production under extreme environmental conditions by alkali-halo-thermophilic bacteria from Saudi Arabia’, Saudi Journal of Biological Sciences, vol. 22, no. 4, pp. 466-475, https://doi.org/10.1016/j.sjbs.2014.11.018
Geetha, SJ, Banat, IM & Joshi, SJ 2018, ‘Biosurfactants: production and potential applications in microbial enhanced oil recovery (meor)’, Biocatalysis and Agricultural Biotechnology, vol. 14, pp. 23-32, https://doi.org/10.1016/j.bcab.2018.01.010
Grötzinger, SW, Schöckel, L, Forchhammer, K & Pleiss, J 2018, ‘Identification and experimental characterization of an extremophilic brine pool alcohol dehydrogenase from single amplified genomes’, ACS Chemical Biology, vol. 13, no. 1, pp. 161-170, https://doi.org/10.1021/acschembio.7b00792
Harrington, LB, Paez-Espino, D, Staahl, BT, Chen, JS, Ma, E, Kyrpides, NC & Doudna, JA 2017, ‘A thermostable Cas9 with increased lifetime in human plasma’, Nature Communications, vol. 8, no. 1, https://doi.org/10.1038/s41467-017-01408-4
Husnawati, Sahlan, M, Cahyana, AH, Maulana, T & Kholisoh, NA 2023, ‘Investigation of Chemical Compounds from Phomopsis Extract as Anti-Breast Cancer Using LC-MS/MS Analysis, Molecular Docking, and Molecular Dynamic Simulations’, International Journal of Technology, vol. 14, no. 7, pp. 1476-1486. Available at: https://doi.org/10.14716/ijtech.v14i7.6696
Jimoh, AA, Lin, J, Tian, Y, Qiu, X, Du, L & Zhao, X 2021, ‘Development and Genetic Engineering of Hyper-Producing Microbial Strains for Improved Synthesis of Biosurfactants’, Molecular Biotechnology, vol. 63, pp. 267-288, https://doi.org/10.1007/s12033-021-00302-1
Karlson, CKS, Lee, S, Siqueira, JE & Park, YS 2021, ‘CRISPR/dCas9-based systems: Mechanisms and applications in plant sciences’, Plants, vol. 10, no. 10, pp. 1-22, https://doi.org/10.3390/plants10102055
Kitchen, DB, Decornez, H, Furr, JR & Bajorath, J 2004, ‘Docking and scoring in virtual screening for drug discovery: Methods and applications’, Nature Reviews Drug Discovery, vol. 3, no. 11, pp. 935-949, https://doi.org/10.1038/nrd1549
Lischer, K., Tansil, K.P., Ginting, M.J., Sahlan, M., Wijanarko, A. & Yohda, M 2020,‘Cloning of DNA Polymerase I Geobacillus thermoleovorans SGAir0734 from a batu kuwung hot spring in escherichia coli‘, International Journal of Technology, vol. 11 no. 5, pp. 921-930 https://doi.org/10.14716/ijtech.v11i5.4311
Makarova, KS, Wolf, YI, Alkhnbashi, OS, Costa, F, Shah, SA, Saunders, SJ, Barrangou, R & Koonin, EV 2015, ‘An updated evolutionary classification of CRISPR-Cas systems’, Nature Reviews Microbiology, vol. 13, no. 11, pp. 722-736, https://doi.org/10.1038/nrmicro3569
Matson, AW, Spina, BJ, Bess, KR, Long, BL & Zhang, H 2019, ‘Optimizing sgRNA length to improve target specificity and efficiency for the GGTA1 gene using the CRISPR/Cas9 gene editing system’, PLoS ONE, vol. 14, no. 12, https://doi.org/10.1371/journal.pone.0226107
Memarpoor-Yazdi, M, Karbalaei-Heidari, HR & Khajeh, K 2017, ‘Production of the renewable extremophile lipase: Valuable biocatalyst with potential usage in food industry’, Food and Bioproducts Processing, vol. 102, pp. 153-166, https://doi.org/10.1016/j.fbp.2016.12.015
Meng, XY, Zhang, HX, Mezei, M & Cui, M 2011, ‘Molecular docking: A powerful approach for structure-based drug discovery’, Current Computer-Aided Drug Design, vol. 7, no. 2, pp. 146-157, https://doi.org/10.2174/157340911795677602
Mengstie, MA & Wondimu, BZ 2021, ‘Mechanism and applications of CRISPR/Cas-9-mediated genome editing’, Biologics: Targets and Therapy, vol. 15, pp. 353-361, https://doi.org/10.2147/BTT.S326422
Mordor Intelligence (n.d.), Biosurfactants Market Size & Share Analysis - Growth Trends & Forecasts (2024 - 2029), https://www.mordorintelligence.com/industry-reports/biosurfactants-market
Mougiakos, I, Bosma, EF, de Vos, WM, van der Oost, J & Schaap, PJ 2017, ‘Efficient genome editing of a facultative thermophile using mesophilic spCas9’, ACS Synthetic Biology, vol. 6, no. 5, pp. 849-861, https://doi.org/10.1021/acssynbio.6b00339
Mougiakos, I, Mohanraju, P, Bosma, EF, Gussak, A, Brinkman, RB & van der Oost, J 2017, ‘Characterizing a thermostable Cas9 for bacterial genome editing and silencing’, Nature Communications, vol. 8, no. 1, https://doi.org/10.1038/s41467-017-01591-4
Nelson, DL & Cox, MM 2001, Principles of Biochemistry, https://doi.org/10.1002/bmb.2005.494033010419
Pramayuditya, M 2023, Molecular Docking Rancangan sgRNA Variasi Panjang Spacer Repeat dan TracrRNA untuk Produksi CRISPR-Cas9 dan dCas9 Geobacillus kaustophilus, Final Thesis, Universitas Indonesia, https://lib.ui.ac.id/m/detail.jsp?id=9999920524491&lokasi=lokal
Qin, Q, Ploux, O, Friehs, K, Dufresne, A & Voget, M 2018, ‘CRISPR/Cas9 editing genome of extremophile Halomonas spp.’, Metabolic Engineering, vol. 47, pp. 219-229, https://doi.org/10.1016/j.ymben.2018.03.018
Roda, FAT & Bassam, TY 2017, ‘Halo-thermophilic bacteria and heterocyst cyanobacteria found adjacent to halophytes at Sabkhas, Qatar: Preliminary study and possible roles’, African Journal of Microbiology Research, vol. 11, no. 34, pp. 1346-1354, https://doi.org/10.5897/ajmr2017.8648
Rodrigues, CJC, Carvalho, LRS, Barreto, LM, Almeida, DS & Goulart, SM 2017, ‘Cultivation-based strategies to find efficient marine biocatalysts’, Biotechnology Journal, vol. 12, no. 7, https://doi.org/10.1002/biot.201700036
Sahlan, M, Sari, MA, Ramadhani, T, Suprihadi, D, Lischer, K & Setiawan, AD 2020, ‘Molecular Docking of South Sulawesi Propolis against Fructose 1,6-Bisphosphatase as a Type 2 Diabetes Mellitus Drug’, International Journal of Technology, vol. 11, no. 5, pp. 910-920, https://doi.org/10.14716/ijtech.v11i5.4332
Sahlan, M, Sari, MA, Ramadhani, T, Wibowo, H, Mahdiana, IT & Fitri, Y 2023, ‘In Silico Identification of Propolis Compounds Potential as COVID-19 Drug Candidates Against SARS-CoV-2 Spike Protein’, International Journal of Technology, vol. 14, no. 2, pp. 387-398, https://doi.org/10.14716/ijtech.v14i2.5052
Sarkar, A & Kellogg, GE 2010, ‘Hydrophobicity-Shake Flasks, Protein Folding and Drug Discovery’, Current Topics in Medicinal Chemistry, vol. 10, no. 1, pp. 67-83, https://doi.org/10.2174/156802610790232233
Sarzynska, J, Popenda, M, Antczak, M. & Szachniuk, M 2023 ‘RNA tertiary structure prediction using RNAComposer in CASP15‘, PROTEINS: Structure, Function, and Bioinformatics, vol. 91, no 12, pp. 1790-1799 https://doi.org/10.1002/prot.26578
Shibulal, B, Babu, S & Singh, A 2014, ‘Microbial enhanced heavy oil recovery by the aid of inhabitant spore-forming bacteria: An insight review’, The Scientific World Journal, https://doi.org/10.1155/2014/309159
Sun, W, Yang, J, Wu, J, Wang, H, Wang, W & Yan, Y 2019, ‘Structures of Neisseria meningitidis Cas9 Complexes in Catalytically Poised and Anti-CRISPR-Inhibited States’, Molecular Cell, vol. 76, no. 6, pp. 938-952.e5, https://doi.org/10.1016/j.molcel.2019.09.025
Uppada, V, Gokara, M & Rasineni, GK 2018, ‘Diagnosis and therapy with CRISPR advanced CRISPR based tools for point of care diagnostics and early therapies’, Gene, vol. 676, pp. 65-76, https://doi.org/10.1016/j.gene.2018.02.066
Yan, Y, Tao, H, He, J, Zhao, Y, Yang, L & Zhang, Y 2020, ‘The HDOCK server for integrated protein-protein docking’, Nature Protocols, vol. 15, no. 5, pp. 1829-1852, https://doi.org/10.1038/s41596-020-0312-x
Yang, J & Zhang, Y 2015, ‘I-TASSER server: New development for protein structure and function predictions’, Nucleic Acids Research, vol. 43, no. W1, pp. W174-W181, https://doi.org/10.1093/nar/gkv342
Zheng, W, Zhang, C, Pearce, R, Bell, EW, Zhang, G & Zhang, Y 2021, ‘Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations’, Cell Reports Methods, vol. 1, no. 3, p. 100014, https://doi.org/10.1016/j.crmeth.2021.100014
Zhou, X, Zheng, W, Li, Y, Pearce, R, Zhang, C, Bell, EW, Zhang, G & Zhang, Y 2022, ‘I-TASSER-MTD: a deep-learning-based platform for multi-domain protein structure and function prediction’, Nature Protocols, vol. 17, pp. 2326-2353, https://doi.org/10.1038/s41596-022-00728-0